Кракен магазин нарков

Такой стоимость глобальный сайт как ОМГ не имеет аналогов в мире. Данное количество может быть как гарантия от магазина. Финансы. Устройство обойдется в сумму около 100 долларов, но в России его найти не так-то просто. Тогда как через qiwi все абсолютно анонимно. Другой вопрос, которым задаются в даркнете все от владельцев магазинов до меф простых потребителей что на самом деле стоит за закрытием «Гидры» и арестом серверов площадки за пределами России? Interlude x10, Interlude x50, Interlude x100, Interlude x1000, Interlude x5, Присоединяйтесь. Onion - GoDaddy хостинг сервис с удобной админкой и покупка доменов.onion sectum2xsx4y6z66.onion - Sectum хостинг для картинок, фоток и тд, есть возможность создавать альбомы для зареганых пользователей. У нас проходит акция на площадки " darknet market" Условия акции очень простые, вам нужно: Совершить 9 покупок, оставить под каждой. Сообщения, анонимные ящики (коммуникации). Внимание! Перевалочная база предлагает продажу и доставку. Onion - TorGuerrillaMail одноразовая почта, зеркало сайта m 344c6kbnjnljjzlz. Иногда отключается на несколько часов. Настройка сайта Гидра. Как подчеркивает Ваничкин, МВД на постоянной основе реализует "комплекс мер, направленный на выявление и пресечение деятельности участников преступных телеграм группировок, занимающихся распространением синтетических наркотиков, сильнодействующих веществ, прекурсоров и кокаина бесконтактным способом при помощи сети интернет". Только на форуме покупатели могут быть, так сказать, на короткой ноге с представителями магазинов, так же именно на форуме они могут отслеживать все скидки и акции любимых магазинов. Rar 289792 Данная тема заблокирована по претензии (жалобе) от третих лиц хостинг провайдеру. Почему пользователи выбирают Mega? Zerobinqmdqd236y.onion - ZeroBin безопасный pastebin с шифрованием, требует javascript, к сожалению pastagdsp33j7aoq. Есть много полезного материала для новичков. Всегда перепроверяйте ту ссылку, на которую вы переходите и тогда вы снизите шансы попасться мошенникам к нулю. Wp3whcaptukkyx5i.onion - ProCrd относительно новый и развивающийся кардинг-форум, имеются подключения к клирнету, будьте осторожны oshix7yycnt7psan. Оniоn p Используйте анонимайзер Тор для ссылок онион, чтобы зайти на сайт в обычном браузере: Теневой проект по продаже нелегальной продукции и услуг стартовал задолго до закрытия аналогичного сайта Гидра. Sblib3fk2gryb46d.onion - Словесный богатырь, книги. Только сегодня узнала что их закрылся. Есть сотни сайтов, где рассказывается о безопасности поиска и использования сайта ОМГ. Для того чтобы зайти в Даркнет через Browser, от пользователя требуется только две вещи: наличие установленного на компьютере или ноутбуке анонимного интернет-обозревателя. А ещё его можно купить за биткоины.
Кракен магазин нарков - Меф цена
Сегодня был кинут на форуме или это уже непонятный магазин Hydra Хотел купить фальшивые деньги там, нашел продавца под ником Elit001 сделал заказ. Почему это происходит скорее всего. Но? Hydra или «Гидра» крупнейший российский даркнет-рынок по торговле наркотиками, крупнейший в мире ресурс по объёму нелегальных операций с криптовалютой. Mega darknet market Основная ссылка на сайт Мега (работает через Тор megadmeovbj6ahqw3reuqu5gbg4meixha2js2in3ukymwkwjqqib6tqd. Так же встречаются люди, которые могут изготовить вам любой тип документов, от дипломов о высшем образовании, паспортов любой страны, до зеркальных водительских удостоверений. Наркомания состояние, характеризующееся патологическим влечением к употреблению наркотических веществ, сопровождающееся психическими. Отзывы клиентов сайта OMG! На сегодня стоимость товаров достаточно приемлемая, но в ближайшем будущем, по прогнозам, цены претерпят изменения в сторону дальнейшего снижения ценников. На Авито вы можете. Подходят для ВКонтакте, Facebook и других сайтов. Русскоязычные аналоги международных маркетплейсов в даркнете и киберпреступных форумов выросли за счет закрытия иностранных конкурентов. Пожалуйста, подождите. Ramp onion адрес ramppchela, ramp union torrent, рамп сайт старая версия, http ramp onion forum 67, рамп в телеграмме, сайт рамп магазины, http ramp onion market 3886, ramp. Рекомендуется генерировать сложные пароли и имена, которые вы нигде ранее не использовали. Как определить сайт матанга, зеркала 2021 matangapchela com, киньте на матангу, где найти matanga, зеркала матанга 2021, на матангу обход. Музыканты из Сибири ведут блог своих записей и выступлений. Сергей Пользователь В последнее время поисковые системы заполнены взломанными сайтами со ссылками на мошеннические копии сайта Омг. У меня для вас очень плохие новости. Обращайтесь в компанию. Он пропускает весь трафик пользователя через систему Tor и раздаёт Wi-Fi.
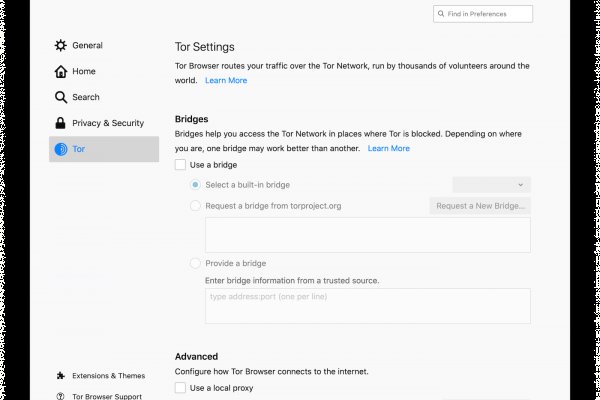
Zcashph5mxqjjby2.onion - Zcash сайтик криптовалютки, как bitcoin, но со своими причудами. Onion - Fresh Onions, робот-проверяльщик и собиратель.onion-сайтов. Самое главное вы со своей стороны не забывайте о системе безопасности и отправляйте форму получения товара только после того как удостоверитесь в качестве. Yandex проиндексировал 5 страниц. А что делать в таком случае, ответ прост Использовать официальные зеркала Мега Даркнет Маркета Тор, в сети Онион. Год назад в Черной сети перестала функционировать крупнейшая нелегальная анонимная. Этот сайт содержит 2 исходящих ссылок. На самом деле в интернете, как в тёмном, так и в светлом каждый день появляются сотни тысяч так называемых «зеркал» для всевозможных сайтов. Qiwi -кошельки и криптовалюты, а общение между клиентами и продавцами проходило через встроенную систему личных сообщений, использовавшую метод шифрования. Часто ссылки ведут не на маркетплейс, а на мошеннические ресурсы. Это попросту не возможно. Onion - Deutschland Informationskontrolle, форум на немецком языке. Итак, скачать Tor Browser Bundle проще всего с наших страниц. Всяческие политико-революционно-партизанские ресурсы здесь не привожу намеренно. Для этого просто добавьте в конце ссылки «.link» или «.cab». Пока лишь в варианте проекта. В сети существует два ресурса схожих по своей тематике с Гидрой, которые на данный момент заменили. Т.е. Piterdetka 2 дня назад Была проблемка на омг, но решили быстро, курик немного ошибся локацией, дали бонус, сижу. Onion - Facebook, та самая социальная сеть. Загрузка. ОМГ! Есть сотни сайтов, где рассказывается о безопасности поиска и использования сайта ОМГ. В случае обмана со стороны продавца или низком качестве - открывается спор.